16.11.2015
НЕКОТОРЫЕ ДИСКУССИОННЫЕ ВОПРОСЫ СТРАТИГРАФИИ И ПАЛЕОГЕОГРАФИИ РАННЕГО АНТРОПОГЕНА
Рубрика: БиологияАвтор: admin
Фауна млекопитающих раннеантропогенового времени прошла в своем развитии ряд этапов.
Смена одного комплекса другим обязана не только общему процессу эволюции различных групп его составляющих, но и изменениям среды обитания.
Восточная Европа входит в ту часть Палеарктической области, которая в четвертичное время претерпела значительное похолодание климата.
Млекопитающие раннего антропогена Восточной Европы дают представление о переходной эпохе, на протяжении которой формировалась фауна, предшествующая развитию на этой территории фауны времени максимального оледенения.
Находясь на стыке двух фаунистических провинций (или подобластей — средиземноморской и центрально-азиатской), фауна юга Европейской части СССР ощущала влияние как с запада, так и с востока.
Изучение млекопитающих эоплейстоцена и нижнего плейстоцена позволило выявить основные этапы развития фауны.
Наблюдаемые в ней изменения состава и перегруппировки позволяют высказать некоторые соображения по ряду дискуссионных проблем четвертичной геологии (древние оледенения, палеогеографические реконструкции, нижняя граница антропогена, объем эоплейстоцена, время и пути прохорезов крупных млекопитающих).
Данные по палеоэкологии млекопитающих широко используются в палеогеографических построениях для территории Европы четвертичного времени. Исходя из фактов истории развития фауны, полигляциалисты доказывают существование нескольких ледниковых эпох.
Моногляциалисты считают, что было одно оледенение с несколькими стадиями, начало которого отвечает максимальному (рисскому) оледенению Западной Европы (Громов, 1948 и др.).
Антигляциолисты приходят к выводу, что масштабы четвертичных оледенений колоссально завышены, и считают, что покровных оледенений на территории Восточной Европы не было (Пидопличко, 1951 и др.).
Вопрос об оледенениях с точки зрения палеонтологических данных (или, вернее, сточки зрения изучения развития фауны млекопитающих) не так прост, как кажется на первый взгляд. Нередко приходится встречаться с такими представлениями: межледниковья связывают с теплой фауной, оледенения — с холодной.
Предполагают, что по мере смены этих эпох меняются и фауны, т.е. имеется многократное повторение «теплых» и «холодных» фаун. На первый взгляд такие представления очень логичны, но, к сожалению, они не отражают действительной истории развития фауны и смены ее комплексов, которая происходила на протяжении четвертичного периода.
В настоящее время мы имеем еще очень слабое представление об арктических комплексах древних ледниковых эпох. Современная арктическая фауна сохранила облик фауны последнего оледенения (вюрмского — в западноевропейской терминологии или осташковского и калининского — в схеме для Восточной Европы).
Отложения этой последней ледниковой эпохи довольно хорошо сохранились, и имеется много местонахождений фауны, которые дают представление о ее составе в то время. Мы знаем, что фауна последней ледниковой эпохи отступила к северу и многие ее элементы продолжают жить сейчас в приполярных и заполярных областях.
Если же мы будем рассматривать фауны млекопитающих времени более древних оледенений, чем верхнеплейстоценовое, то в этом случае получим несколько другую картину. Вся сложность в том, что в то время имелись иные комплексы, резко отличные от холодоустойчивых комплексов верхнего плейстоцена.
Прежде всего следует учесть, что местонахождений фауны древних эпох оледенений значительно меньше, так как их отложения подверглись сильному разрушению и на Европейской части СССР сохранились в основном в переуглублениях. По мнению большинства геологов, следы миндельского оледенения установлены в ряде мест на территории Западной и Центральной Европы.
Но не всюду на территории северной Евразии находят достоверные следы покровного оледенения этого времени. Так, оно отрицается для многих районов Центральной Азии (Равский, 1972). Однако уже появились материалы, позволяющие видеть весьма холодоустойчивый комплекс фауны млекопитающих (Шер, 1971), обитавший на северо-востоке Азии в эпоху, отвечающую миндельскому оледенению Европы.
В одно и то же время в различных районах естественно существовали и разные условия. Приспосабливаясь к обстановке развивающейся ледниковой эпохи, фауна не во всех районах будет изменяться одинаково. Хорошо известно, что различные виды животных были в разной степени восприимчивы к климатическим условиям.
Представители многих групп животных, сходные по ряду анатомических и адаптивных признаков с обитателями теплых районов, могли существовать в иных условиях (например, слоны и носороги). Многие животные того отдаленного времени обладали широкой экологической пластичностью.
При отступании ледника часть фауны, обитавшей близ ледникового щита, следовала за отступающим его краем. Другая же часть оставалась на месте, становясь частью нового формирующегося комплекса. Этот новый комплекс внеледниковой фауны формировался в большой мере за счет пришельцев из других районов. Получалось смешение форм неледниковых и бывших ледниковых.
Это перемешивание в какой—то мере может послужить объяснением того, что фауны млекопитающих последовательных ледниковых и межледниковых эпох никогда не повторяют точно одна другую, хотя изменения климата носят примерно один и тот же характер.
Конечно, здесь всегда следует учитывать и общую эволюцию, которая несколько меняла облик фауны, не допуская повторения.
Местонахождения раннечетвертичной фауны млекопитающих территории юга Европейской части СССР располагаются в зоне, в которой не найдены бесспорные ледниковые отложения, Фауна этой зоны не дает прямых доказательств воздействия ледниковых процессов.
Однако постепенное исчезновение от комплекса к комплексу теплолюбивых элементов из состава фауны свидетельствует об общем прогрессирующем изменении климата в сторону похолодания.
По фауне млекопитающих эоплейстоцена рассматриваемой территории на данном уровне знаний нельзя решить вопрос: были ли это крупные периодические колебания климата или происходило более или менее ровное усиление похолодания. Никаких следов повторных миграций теплолюбивых элементов не найдено.
В период существования фауны молдавского комплекса на территории юга Европейской части СССР и прилежащих районов существовал климат, отдаленно напоминавший климат субтропического типа: сухое и довольно жаркое лето и весьма влажная зима.
Основная масса животных молдавского комплекса тяготела к жизни в условиях лесостепи или саваннового ландшафта (ананкоидные мастодонты, лошади, многие полевки, пищухи и др.). Однако в фауне присутствовал целый ряд форм, основным биотопом которых является лес (мастодонты Борсона, обезьяны, тапиры, свиньи медведи, белки, сони).
Это позволяет допустить существование -больших лесных массивов в западных районах и на территории Северного Кавказа. В отличие от западноевропейских фаун рассматриваемого времени в молдавском комплексе присутствуют верблюды. Огромные скопления их костей найдены в фауне Одесских катакомб, в небольших количествах остатки встречены в местонахождениях долин Большой Сальчи и Кагула (в. основном в южных пунктах).
В долине Кучургана находки верблюдов единичны. Видимо, только причерноморская полоса в своей западной части имела открытые ландшафты, где существовали условия жаркой степи. Безусловно, сравнительно с территорией Западной и Центральной Европы залесенность восточноевропейских районов была выражена слабее.
Ни фауна млекопитающих, ни осадки, с которыми связаны изученные местонахождения молдавского комплекса, еще не дают сведений об изменениях климата в сторону похолодания. Большинство исследователей единодушны в мнении, что климат этого времени был жарким и переменно влажным (Молявко, 1960; Веклич, 1961; Никифорова и др., 1965; Ренгартен, Константинова, 1965; Ренгартен, 1971; и др.).
Заметные изменения климата в сторону общей аридизации и нарастающего похолодания начинаются во время существования ранней стадии развития хапровского комплекса.
На территории юга Европейской части СССР уже на переходе от молдавского комплекса к следующему, хапровскому, исчезает ряд теплолюбивых животных, таких, как жирафы, тапиры, мунтжаки, динотерии, бегемоты, агриотерии, амфиционы. Наблюдается сокращение ареалов мастодонтов, махайродов, гиппарионов, обезьян.
На протяжении развития хапровского комплекса идет приспособление фауны к обитанию в условиях открытых ландшафтов. Среди крупных млекопитающих доминируют группы животных с гипсодонтными зубами, как наиболее приспособленные к питанию жесткой растительностью степей.
Это — эласмотерии, слоны, лошади, верблюды. Развитие цемента на зубах корнезубых полевок тоже, по-видидимому, является своеобразным приспособлением к обитанию в условиях довольно сухих ландшафтов. В фауне этого времени встречаются страусы, существование которых обычно рассматривают как неоспоримое доказательство теплой или весьма теплой климатической обстановки (Бурчак-Абрамович, Конькова, 1967).
Несмотря на усилившееся остепнение территории кга Европейской части СССР, нельзя утверждать, что это было время господства только степных форм ландшафта.
Существовали также залесенные массивы (особенно в приустьевых участках рек). В хапровском комплексе еще встречаются обезьяны, появляются лоси, нередки находки медведей, обильно представлены олени. Эти животные обычно связаны с лесными биотопами.
Климат южных районов Европейской части СССР в хапровское время был довольно теплым. Однако изменения состава комплекса на протяжении его развития (быстрые эволюционные изменения архидискодонтных слонов, исчезновение обезьян и гребнезубых мастодонтов, перестройка фауны грызунов) позволяют предполагать, что процесс нарастания похолодания прогрессировал.
В период существования таманского комплекса продолжается дальнейшее развитие фауны степного типа и шлифовка ее приспособлений к обитанию в условиях нарастающего похолодания и увлажнения. Преобладают в фауне крупные формы лошадей, эласмотерии, зубры, олени (сложнорогие, благородные, большерогие), крупные лоси, кабаны, антилопы, южные слоны (высокоразвитая поздняя форма).
Последние овернские мастодонты полностью исчезают в таманском комплексе. Преобладающей группой среди грызунов таманской фауны продолжают оставаться полевки родов Lagurodon и Allophaimys. Мимомисная группа испытывает заметное угнетение; появляются полевки рода Clethrionomys.
Судя по общему облику таманской фауны, на территории юга Восточной Европы господствовали те же ландшафты, что и в предшествующее время. Однако обилие оленей, присутствие лосей и кабанов указывают на значительную залесенность, в частности — для равнинных районов территории северного Предкавказья.
Фауна грызунов Ногайского местонахождения (средний горизонт Ногайского разреза) также свидетельствует об увеличении площадей приречных лесов и о некотором увлажнении климата.
Фауны Западной Европы, более или менее синхронные таманской, существенно лесные (первые настоящие локсодонтные слоны, много оленей, бегемоты, кое—где даже мартышкообразные обезьяны). Это фауны кромерского типа или, как иногда их называют, фауны конечного виллафранка.
В работах западноевропейских геологов и палеонтологов (Kurten, 1963; Bourdier е. а., 1969; Chaline, 1972) имеются данные о присутствии холодоустойчивых элементов в фауне виллафранка. На территории Франции в отложениях, отвечающих конечному виллафранку, вместе с холодной фауной моллюсков обнаружены костные остатки Dicrostonys sp (карьер Журден, Амьен) и Lemmus lemmus (район Парижа).
Первое местонахождение связано с лёссом позднего гюнца, второе — с гюнц—минделем. Комплекс моллюсков из района Амьена указывает на умеренно—влажную обстановку, несколько более холодную, чем современная.
Тираспольский комплекс хорошо отличается от хапровско—таманской фауны. Это пока первая фауна на территории Восточной Европы, состав которой позволяет делать выводы в пользу сильного похолодания, близкого по своим размерам к эпохе максимального оледенения. Сейчас геологические материалы дают все больше и больше оснований думать о значительности миндельского оледенения.
Моренные отложения доднепровского времени хорошо известны в Белоруссии (Цапенко, 1960 и др.). На территории Украины в котловане Каневской ГЭС и в скважинах в районе ее строительства вскрыта мощная толща ледниковых отложений ц ленточных глин, имеющих доднепровский возраст (Ромоданова и др., 1969).
Эта серия отложений сохранилась только в переуглубленных участках долины Днепра (Ромоданова и др., 1969). Морена Каневского котлована показывает, как далеко на юг простиралось миндельское оледенение. Его влияние не могло не сказаться на фауне млекопитающих, населявших северное Причерноморье.
По экологическому составу тираспольская фауна в целом очень разнообразна. Наряду с лесостепными слонами—архидискодонтами (слон Вюста), этрусским носорогом, зубрами и другими, в ее состав входят обитатели лесов (разнообразные олени, лоси, косули, носороги Мерка, медведи), а также жители относительно сухих полустепных стаций ( стройные, быстро бегающие антилопы-понтороги, лошади, полуослы, верблюды, на востоке рассматриваемого района—эласмотерии).
Обилие лосей и разнообразие оленей позволяют думать, что на западе нередко были развиты участки ландшафта болотистого и кустарникового леса, на востоке — сухого лесостепного типа.
Большинство местонахождений тираспольского комплекса находится в южной зоне Европейской части СССР, где нет настоящих ледниковых отложений миндельского времени. Естественно, в этих местонахождениях не найдены типичные элементы «ледниковой» фауны. Тем не менее анализ родового состава тираспольского комплекса позволяет считать его холодоустойчивым.
В фауне этого времени присутствуют преимущественно те животные, прямые потомки которых довольно хорошо пережили два последующих оледенения (первое из которых большинством геологов сейчас считается масимальным). Это — быки, давшие начало Bison priscus; благородные и большерогие олени, лоси, лошади кабаллоидной группы, слоны Вюста (от которых, по мнению большинства палеонтологов, развивается мамонтовая линия).
В тираспольское время начинается формирование так называемой пещерной фауны хищников. В это же время появляются первые лемминги. Все это позволяет думать, что на начальном.этапе развития миндельского оледенения начался отбор групп млекопитающих,шедший, образно выражаясь, через «сито холода». Формирование тираспоЛьской фауны может быть понято как реакция на значительное похолодание.
Пока еще нельзя четко выявить основные причины изменения состава фауны. Для каждой группы млекопитающих это свои специфические причины (усиление залесенности, возрастание толщины снежного покрова, повышение влажности, смещение цикла воспроизводства и др.), в той или иной мере связанные с общим изменением климатической обстановки.
Второй крупной дискуссионной проблемой четвертичной геологии является вопрос о нижней границе антропогена и об объеме эоплейстоцена (в частности, о его верхнем пределе). При обсуждении этого вопроса большое значение придается фауне млекопитающих и ее изменениям.
Большинство геологов положительно оценивают рекомендации XVIII сессии Международного геологического конгресса в Лондоне (1948 г.) относительно снижения границы под калабрий. Прошедший Международный коллоквиум по проблеме нижней границы антропогена (июнь 1972 г.) констатировал, что имеют место три заметных рубежа.
Большинство участников высказалось за проведение границы под калабрием и его континентальным аналогом — верхним виллафранком. Это решение’ было подтверждено последней сессией Геологического конгресса (сентябрь 1972 г.).
Большинство палеонтологов-маммологов признают необходимость снижения нижней границы. На прилагаемой табл. 13 отражены мнения исследователей, которые рассматривают вопрос об этой границе в Восточной Европе с точки зрения развития фауны млекопитающих. Единодушия нет. Вся сложность состоит в том, что неясно, каков объем виллафранка и каковы его аналоги за пределами Италии.
По поводу объема виллафранка и его деления нет единого мнения и у западноевропейских геологов. Если же учесть еще одно замечание Конгресса 1948 г., сделанное в указанной рекомендации по поводу нижней границы, то ее проводить следует по первому появлению в фауне виллафранка слонов, быков и настоящих лошадей.
Тогда в Восточной Европе нижнюю границу правомерно провести под отложениями с молдавским комплексом (под акчагылом), в составе фауны которого известны самые древние находки остатков Equus и Archidiskodon. Если же проводить границу по времени широкого распространения этих родов (по расцвету фауны), то тогда ее логично провести под отложениями, содержащими фауну поздней стадии хапровского комплекса.
Изучение раннеантропогеновой фауны млекопитающих позволило выявить ряд особенностей в ее развитии. Эти результаты исследования могут быть полезны при работах по уточнению положения нижней границы антропогена в Восточной , Европе и установлению восточноевропейских аналогов виллафранка юга Западной Европы.
В истории развития раннечетвертичной фауны млекопитающих Восточной Европы можно подметить три крупных рубежа как с точки зрения ее эволюционных превращений, так и в смысле появления мигрантов. Первое резкое изменение приходится на начало молдавского комплекса.
В отличие от позднегиппарионовой фауны появляется ряд новых элементов (однопалые лошади, первые архидискодонтные слоны, олени рода Cervus, мастодонты рода Anancus, обезьяны-долихопитеки и др.). Наблю дается заметная перегруппировка среди мастодонтов. Появившийся род Anancus в кратчайшее время получает широкое распространение в зоне теплых широт Старого Света.
Второй рубеж имел место между молдавским и хапровским комплексами (быстр исчезает ряд реликтовых форм и происходит повсеместное распространение слонов и лошадей). Третий рубеж отчетливо устанавливается на переходе между таманским и тираспольским комплексами (о характере изменений в составе фауны речь шла выше).
Таманский комплекс тесно связан с хапровской фауной, представляя собой завершающее звено ее развития.
Тираспольский комплекс — это ранний этап в развитии фауны плейстоцена, приведший в конечном итоге к формированию современной фауны Восточной Европы.
Существенное различие этих двух комплексов позволяет говорить о необходимости пересмотра вопроса о верхней границе эоплейстоцена.
Вероятно, правильнее вернуться к тому объему эоплейстоцена, который был предложен В.И. Громовым в 1957 г. К.В. Никифорова (1969) также считает более целесообразным проведение границы между эоплейстоценом и плейстоценом под отложениями с тираспольским комплексом.
Процесс формирования комплексов и их смены весьма сложен. Если в миоцене для формирования гиппарионовой фауны Евразии огромное значение имел африканский центр развития фауны (Schlosser, 1903). То картина формирования ранне-четвертичной фауны несколько иная.
Доминирующую роль в процессе ее становления играл азиатский центр развития фауны (Алексеева, 1964а, 19666). Проникшие сюда азиатские элементы вошли в состав существовавшей здесь фауны, вызвав ее перегруппировку (Алексеева, 1973а,б).
На протяжении этого времени имели место две фаунистические волны. Появление первой волны новых элементов фауны относится к раннему эоплейстоцену (предвиллафранкское время). В это время на территории Европы появился целый ряд новых родов, непосредственно не связанных с предшествующей фауной.
Появление таких родов в Европе не дает оснований говорить о их времени возникновения. Л.Ш.Давиташвили (1970) рекомендует геологам-палеонтологам быть осторожными в своих выводах, так как в подобном случае устанавливается не момент возникновения нового таксона, а появление его на данном, участке. И именно поэтому можно утверждать, что многие роды ( иммигранты или виды-иммигранты) могут служить хорошими биостратиграфическими реперами для корреляции отложений на территории, занятой ими во время экспансии.
Примером может служить род Hip-parion, для распространения которого на территории Европы потребовался довольно короткий промежуток времени (Габуния, 1959).
Что касается эволюции автохтонных видов, то, как правило, для преобразований такого рода требуется значительно больше времени. Изучая виллафранкских оленей, Э. Эйнц (Heintz, 1970) пришел к выводу, что для развития подвида требуется около 500 000 лет.
Эти два примера показывают то большое значение, которое имеют иммигранты при сопоставлениях отложений удаленных территорий. Это ни в какой мере не принижает значение эволюционных преобразований автохтонной фауны, которые составляют основу местных биостратиграфических схем.
В течение эоплейстоцена и в начале плейстоцена на территории восточноевропейской области и в сопредельных районах появилась целая группа родов млекопитающих (Archidiskodon, Palaeoloxodon, Equus, Elasmotherium, Leptobos, Bison, Cervus, Eucladoceros, Alces, Gazellospira, Vulpes, Nyctereutes, Panthera, Crocuta и др.), прародиной которых в основном являлись те или иные районы Центральной и Юго-Восточной Азии.
С территорией Азии связаны группа реннечетвертичных полорогих (быки, антилопы), группа свиней, многие роды оленеобразных (Кришнан, 1954). В сиваликской серии в горизонте докпатан (примерно одновременно понту) уже встречаются такие роды, как Propotamochoerus, Hippopotamus, Cervus, Proleptobos, Taurotragus, Agriotherium, Macaca.
Несколько позже — роды Equus и Sus в татроте, роды Protelephas и Bison — в пинджоре. До сих пор остается нерешенным вопрос о месте формирования ранних слонов и особенно рода Archidiskodon, с которым связывают линию мамонтов плейстоцена. Распространение предковых форм (стегодонов и стеголофодонов) в Юго-Восточной Азии позволяет рассматривать эту часть Палеоарктики как наиболее вероятный район возникновения слонов рода Archidiskodon (Алексеева, 19616). Имеется иная точка зрения, отстаивающая африканское происхождение этой группы (Osborn, 1942; Гарутт, 1965; Maglio, 1970, 1973).
В последнее время появилось много новых сведений из местонахождений Восточной Африки, свидетельствующих о существовании там очень древних форм слонов. В. Малье (Maglio, 1970) дает датировку для первых африканских слонов 5,5 млн. лет.
Следует, однако, отметить, что современные африканские слоны обнаруживают родство с этими древними формами, относящимися к роду Primelephas, а слоны рода Archidiskodon более близко родственны во всех звеньях эволюционной цепи группе азиатских слонов линии Protelephas. Что касается рода Palaeoloxodon, то его азиатское происхождение пока не оспаривается.
Прохорезы в группе непарнопалых тоже имели направление с востока на запад. Лошади рода Equus, проникнув из Северной Америки в Старый Свет, естественно, заселили вначале азиатскую часть материка и уже оттуда распространились на запад до самых крайних точек Европы.
В связи с этим вполне понятно более раннее появление однопалых лошадей на территории СССР (бетекейскии и молдавский комплексы). Область формирования рода Е las mother ium пока не установлена. Ареал этого рода был ограничен Восточной Европой и Казахстаном с прилегающими районами.
Логичнее предположить, что прародиной явились какие-то степные районы азиатской части континента (в Западной Европе достоверных находок этого рода не известно). Некоторое пополнение наблюдается и в группе носорогов. В начале плейстоцена на территории Европы появляется носорог Мерка, которого большинство палеонтологов считают не потомком Dicerorhinus etuscus, а пришельцем из Азии (Azzaroli, 1964b и др.).
Азиатское происхождение большинства групп полорогих не вызывает сомнений (Pilgrim, 1939, 1941, 1947 и др.). И.И. Соколов (1953) высказал мнение, чтр основной район центра их развития и расселения находился в Малой Азии или Аравии. К.К. Флеров (1972) считает, что происхождение рода Bison связано с Восточной Азией.
Самые ранние находки Bos, Bison и Leptobos известны на территории Индии и Китая. В Китае Bison sp. найден вместе с Elephas cf. planifrons (Licent, Trassaert, 1935) в плиоценовых отложениях (условно относимых к среднему плиоцену). В Индии Bison и Bos (Кришнан, 1954) указываются в пинджоре, a Leptobos (Wadia, 1951).— в татроте.
Значительно хуже изучена группа оленей. Изучение виллафранкских оленей Франции и Испании (Heintz, 1970) показало, что в Европе отсутствуют предковые формы этих родов. Сведения о самых ранних находках остатков рода Cervus связаны с док—патанским горизонтом сиваликской серии.
Несколько более раннее появление на территории восточноевропейской области таких родов, как Eucladoceros и Libralces (Alces?), позволяет предполагать, что пути их прохореза имели восточно-западное направление.
По поводу прохорезов хищников раннеантропогеновой фауны утверждать что-либо определенное трудно (в местонахождениях Европейской части СССР очень мало находок). Ж. Вирэ (Viret, 1954) рассматривает род Nyctereutes как азиатского мигранта, пришедшего в Западную Европу в виллафранкское время. Н.К. Верещагин (1971), отмечая факт-особого разнообразия кошек Юго-Восточной Азии, считает, что там—то и был центр их происхождения. Формирование пещерного льва, появившегося в Европе в нижнеплейстоценовое время, этот исследователь связывает с субарктической зоной Палеарктики, а не с миграцией его из Африки в Европу (Рябинин, 1937).
Разбирая вопросы формирования мелких млекопитающих, Ж. Шалин (1972) утверждает, что в конце виллафранка Западную Европу заселяют такие азиатские иммигранты, как роды Allophaiomys, Dicrostonyx, Lagurus, Citellus, Allocricetus, Sicista и др.
Глубокое обновление, отмечаемое в руссильонской фауне Западной Европы, в молдавском комплексе Восточной Европы, в среднесинапской фауне Ближнего Востока, шло не только за счет появления азиатских элементов, В это время имел место и широкий двусторонний обмен между фаунами Евразии и Африки. В результате существовавших тогда сухопутных связей из Африки в Европу прошли ананкоидные мастодонты и даманы.
Если мастодонты рода Anancus получили широчайшее распространение, то даманы заселили лишь самые окраины евразийского континента (их находки известны только на юге Западной Европы и в Закавказье) К. Арамбур (Arambourg, 1969) считал, что современный облик североафриканской фауны (эфиопско-индусский) обязан именно этому времени.
Появившееся недавно сообщение (Jaeger, 1971) о находках зайцеобразных евразиатского рода Prolagus, проникшего в Африку в предвиллафранкское время, подтверждает вывод К.Арабмура относительно существования в этот период тесной связи между фаунами Африки и Евразии.
Проникновение евразиатской фауны в Северную Африку шло как через Закавказье и Ближний Восток, так и через район Гибралтара. Однако путь перехода некоторых животных был несколько иным. Так, прохорез рода Leptobos из Азии в Европу проходил, вероятно, южнее азиатских областей СССР, по территории Ближнего Востока, затем по Северной Африке и Пиренейскому полуострову (рис. 11).
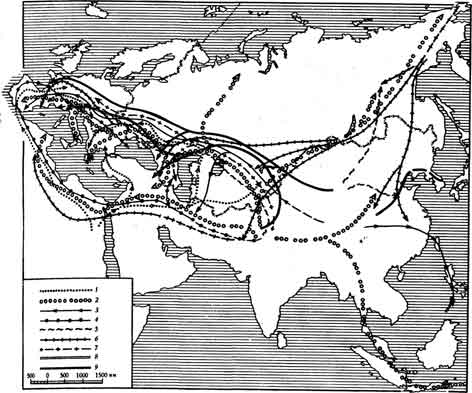 Рис. 11. Предполагаемые пути миграции основных групп крупных млекопитающих
Рис. 11. Предполагаемые пути миграции основных групп крупных млекопитающих
1 — Anancus; 2 — Archidiskodon’, 3 — Palaeoloxodon; 4 — Leptobos; 5 — Bison; 6 — Equus; 7 — Cervus; 8 — Sus и Propotamochoerus; 9 — Nyctereutes
Вторая волна, или, вернее, второй период расселения евразиатской фауны, относится к поствиллафранкскому времени. Это было в основном одностороннее движение (доминирует направление с севера на юг).
В этот период в Северную Африку проникли трогонтериевые слоны, носороги Мерка, быки, олени. Но эти формы не оказали какого-либо заметного влияния на существовавшие там комплексы.
Происшедшее на этом рубеже изменение климата, связанное с похолоданием, явилось наиболее вероятной причиной изоляции фауны Европы от влияния фаун, развивавшихся в то время на территории Северной! Африки и Ближнего Востока.
Вероятно, такая односторонняя миграция может рассматриваться как еще одно косвенное доказательство значительной суровости климата юга Европы, вызвавшей сдвиг ландшафтно-климатических зон и соответствующей им фауны к югу.
Вас также заинтересует это:
Comments (0)
Комментариев нет
Нет комментариев.