20.06.2016
Класс моногеней (Monogenoidea)
Рубрика: ЭволюцияАвтор: admin
Группа моногенетических сосальщиков, по нашему мнению, -правильно возводимая Быховским (1937) в ранг особого класса, равноценного классу Trematodes, в котором сохраняются только дигенетические сосальщики, наряду с многочисленными примерами олигомеризации, дает и некоторые случаи умножения числа гомологичных органов.
Кроме того, в некоторых случаях можно наблюдать у Monogenoidea и начальные этапы олигомеризации, т. е. переход от множественного состояния органов (множественная закладка) к постепенной олигомеризации.
Такую картину дает нам род Tristoma, где мы имеем ряд переходов от Т. integrum (рис. 10, а) с огромным числом кожных шипов до очень небольшого числа этих шипов у Т. coccineum (рис. 10, б). При этом редукции подвергается как число рядов кожных шипов, так и само их количество.
Эктосоматические органы
Прикрепительные аппараты, т. е. крючки и присоски, вообще дают целый ряд примеров изменения их числа, носящих преимущественно характер олигомеризации.
Крючковый аппарат моногеней состоит, как известно, из двух категорий органов: из эмбриональных крючков, вполне развитых уже у личинки и из так называемых „срединных крючьев», нередко появляющихся более поздно.
Быховский признает за крючками большое систематическое и филогенетическое значение. Число эмбриональных крючков у одной группы семейств варьирует от 16 до 14; у другой группы (семейства Octocotylidae и Microcotylidae) имеется 12—10 эмбриональных крючков, притом иной формы, чем у прочих моногеней. Расположение эмбриональных крючков у всех видов вполне правильное (чаще всего в виде венца) и определенное.
Строгая определенность числа крючков и их локализованность заставляют предполагать, что современному состоянию системы эмбриональных крючков предшествовало образование очень многочисленных, неправильно разбросанных по прикрепительному диску, еще более мелких первичных крючочков, или шипиков.
По крайней мере, в пользу этого предположения говорит большая множественность различных кожных дифференцировок (грушевидных органов, чувствительных сенсилл, рабдитных желез и т. д.) у различных турбеллярий.
В таком случае современные эмбриональные крючки представляют собой до известной степени вторичные образования, специализировавшиеся у двух групп моногеней в двух, несколько различных направлениях.
Таким образом, установлению современного типа крючковой формулы предшествовал процесс олигомеризации первичных крючочков.
Быховский подчеркивает большую стабильность числа эмбриональных крючков в обеих группах семейств моногеней. Изменения количества крючков имеют весьма малый диапазон и могут, повидимому, совершаться в обоих направлениях.
По крайней мере, принимая классификацию Быховского, мы находим, что в основном семействе подотряда Dactylogyrinea, а именно Dactylogyridae (рис. 10, в), наблюдается наличие 14 крючков, тогда как у более специализированного семейства Protodactylogyridae имеется 12 крючков, т. е., повидимому, имеет место олигомеризация.
Относительно эволюции числа эмбриональных крючков у некоторых Microcotylidae весьма интересные и единственные в своем роде данные нашел Быховский: во время метаморфоза десяти-крючковой личинки (рис. 10, г) на известной стадии задняя половина прикрепительного диска вместе с 6 эмбриональными крючками автотомирует и отрывается от тела.
Таким образом, происходит явная олигомеризация, и притом по-особому, чрезвычайно-редко встречающемуся типу — посредством автотомии части гомологичных органов (рис. 10, д и ё).
Интерес в вопросе об олигомеризации и полимеризации представляет и судьба оставшихся четырех крючков личинок, микрокотилид. В области этих крючков закладываются четыре первые, а именно самые задние прикрепительные клапаны микрокотилид, причем данные 4 крючка принимают самое непосредственное участие в образовании их хитиноидного аппарата.
Прочие многочисленные клапаны возникают несколько позже и не за счет крючков, а самостоятельно. Отсюда можно сделать следующий существенный вывод: во-первых, все передние клапаны микрокотилид имеют иное происхождение, чем четыре задних и потому не гомологичны им; во-вторых, ввиду всего только что сказанного, образование многочисленных передних клапанов должно рассматриваться не как полимеризация (т. е. умножение числа гомологичных органов), а как множественная закладка новых органов, не гомологичных задним четырем клапанам.
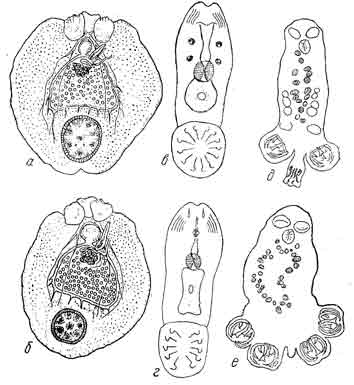
Рис. 10.
a — Tristoma (Capsala) integrum с множеством мелких шипиков кутикулы;
б— Tristoma (Capsala) cocctneum с уменьшенным количеством шипиков кутикулы;
в — расположение эмбриональных крючков у семейства Dactylogyridae (14 крючков);
г — расположение эмбриональных крючков у семейств Осtocotylidae и Microcotylidae— 1° крючков;
d, е.— две стадии метаморфоза личинки Microcotyle:
d — личинка перед самым отрыванием церкомера,
е — личинка вслед за отрыванием церкомера. (Из Быховского).
У некоторых моногеней эмбриональные крючки испытывают олигомеризацию особого рода, построенную на принципе смены функций. Они начинают дифференцироваться на две группы: одну, .сохраняющую первоначальную форму и функцию, другую же, которая начинает функционально заменять или дополнять срединные крючки, принимая иную форму.
Примером может служить Dactylogyrus эркута, у которого одна пара боковых, т. е. эмбриональных крючков, становится очень крупной и получает иную форму, а остальные 12 остаются прежними.

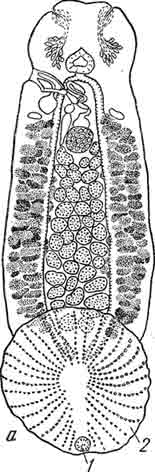
Рис. 11. Справа — типичная форма крючкового аппарата рода Dactylogyrus с двумя поперечными пластинками и четырьмя главными крючками; слева — ближе неопределенный тропический вид Dactytogyrus, у которого один из главных крючков в 10 раз меньше других — тенденция к олигомеризации. (Из Быховского).
Здесь, следовательно, мы имеем в менее сильно выраженном виде тот же процесс смены функций, который в более яркой форме встречается нам в многообразных превращениях нефридиев Рrоtracheata в слюнные железы, анальные железы и т. д.: превращение гомодинамных гомологичных органов в гетеродинамные.
В противоположность эмбриональным крючкам, срединные крючки моногеней во время своей эволюции, по мнению Быховского, испытывают скорее полимеризацию от 2 до 4 и до 6.
Однако и в отношении срединных крючков Быховский отмечает своеобразную олигомеризацию у одного еще не описанного им рода низших Monogenoidea путем прекращения роста одного из 4 имеющихся у данного рода крючков, так что у взрослых червей вместо 4 одинаковых срединных крючков остается 3, а 4-й недоразвитый, хотя и остается, но в 10 раз мельче и иной формы. Такой случай можно рассматривать как не доведенную до конца редукцию одного из крючков (рис. 11).
Возвращаясь к рассмотренным нами ранее мелким прикрепительным клапанам Octocotylidae и Microcotylidae, которые, по Быховскому, служат для позднейшей замены эмбриональных I крючков личинки, мы находим у них прекрасный пример множественной закладки этих образований. Число клапанов варьирует от 8 до 240; только в исключительных случаях оно спускается до 6, 5 и даже 4. Нам кажется, что исходным числом у вышеназванных семейств было 8 клапанов. Формы с 6, 5 и 4 клапанами испытали олигомеризацию. Что касается многоклапанных форм Microcotylidae, то множественность их клапанов зависит, как мы говорили, от того, что клапаны их представляют вторичные образования, возникшие взамен предшествующей их развитию олигомеризации эмбриональных крючков с 10 до 4 (см. стр. 45).
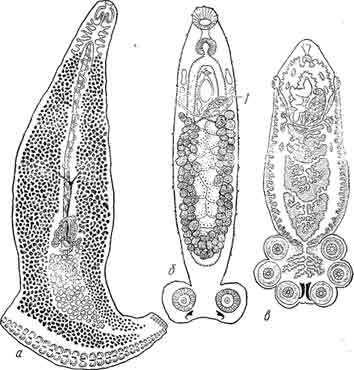
Рис. 12. а — род Heteraxine с асимметричным устройством прикрепительного аппарата: на правой стороне около 30 прикрепительных клапанов, на левой 7; б, в — олигомеризация числа присосок V рода Sphyranura (б) по сравнению с более примитивным родом Polystomum (в). (Из Быховского).
У таких Microcotylidae, как Heteraxine (рис. 12, а), у которых задний конец устроен совершенно асимметрично, на одной стороне задней половины тела имеется несколько десятков более крупных клапанов, на другой же стороне — всего 6 — 7 и притом рудиментарных. Имеется основание думать, что в основе асимметрии у таких форм лежит рудиментация и, далее, олигомеризация числа клапанов на одной стороне тела.
Наконец, несомненно, что число присосок у Sphyranura (всего 2 присоски, рис. 12, б) есть признак вторичный по сравнению с числом присосок у Poly stoma и Polystomoides (6 присосок, рис. 12, в), так как все внутреннее строение говорит о вторичном упрощении организации у Sphyranura (например,вагинальные протоки не имеют выводных отверстий и превращены в своеобразные семеприемники).
 Специальный интерес представляет крючковы й аппарат рода Acanthocotyle (рис. 13), относящегося к семейству Monocotylidae из подотряда Monoopisthocotylinea.
Специальный интерес представляет крючковы й аппарат рода Acanthocotyle (рис. 13), относящегося к семейству Monocotylidae из подотряда Monoopisthocotylinea.
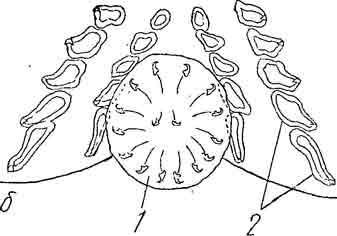
Рис. 13. а —Acanthocotyle с прикрепительным диском при малым увеличении; б— задний конец прикрепительного диска Acanthocotyle при большом увеличении (1 — эмбриональные крючки диска, 2— вторичные крючки на дефинитивном присасывательном диске). (Из Быховского).
У данного рода прикрепительный диск личинки имеет обычное для большинства личинок количествоэмбриональных крючков, а именно 14, и 2 недоразвитых срединных крючка.
Но этот диск имеет лишь провизорный характер и остается миниатюрным, тогда как непосредственно впереди него образуется гораздо более крупный дефинитивный диск. Последний вооружен громадным количеством (сотни) очень мелких вторичных крючочков, расположенных в многочисленные, сходящиеся к центру диска радиальные ряды.
Такая картина толкуется нами следующим образом: личиночный диск, как мы говорили ранее (стр. 45), находится уже на относительно поздней стадии своей эволюции, когда первоначальная множественная закладка эмбриональных крючков в результате олигомеризации свелась к выработке небольшого и определенного числа крючьев.
Дефинитивный диск, очевидно, представляет собою значительно более позднее образование и появляющиеся на нем заново крючки возникают (соответственно с принципом множественной закладки) в очень большом и неопределенном количестве. Поэтому дефинитивный диск Acanthocotyle до известной степени отражает то состояние, в котором некогда находилась крючковая система эмбриональных крючков большинства моногеней.
Глаза.
Вследствие паразитического образа жизни глаза, моногеней вообще склонны к редукции. У одной группы семейстн их 4, у другой 2. Однако у этой второй группы (семейства Dactylogyridae и Monocotylidae) оба глаза склонны к слиянию, т. е. к олигомеризации. При этом слившиеся два глаза сохраняют свои линзочки.
Головные железы.
Быховский считает, что можно рассматривать как один из истинных примеров олигомеризации постепенное сгруппирование протоков головных желез моногеней во все меньшее и меньшее число пучков.
Внешне это отражается на уменьшении числа головных выростов (на которых собраны пучки протоков желез) от 8 к 6, 4 и даже 2.
Процесс этот виден у низших моногеней — Monocotylidae и Dactylogyridae — достаточно четко. В первоначальном своем виде многочисленные протоки желез покрывали передний край головы ровной каймой из многочисленных тонких канальцев.
Вас также заинтересует это:
Comments (0)
Комментариев нет
Нет комментариев.