22.01.2016
Гидроидные медузы
Рубрика: ЭволюцияАвтор: admin
Вопрос о какой-нибудь направленности в изменении числа гомологичных органов во время эволюции гидромедуз находится пока в весьма трудном и даже неразрешимом положении.
Дело в том, что самый ход эволюции этой группы, как взятой в целом, так и в пределах отдельных семейств, остается для нас неясным.
Мы не знаем, происходят ли Trachylina от гидроидных медуз или (к чему склоняется Хайман) эволюция шла в обратном направлении, или же, наконец, и те и другие независимо друг от друга ведут начало от каких-то общих предков (по Кюкенталю).
Такая же неясность царит иногда и в вопросе о родственных отношениях между родами в пределах отдельных семейств.
Органами, которые подвергаются изменениям в числе, у гидроидных медуз являются щупальца, статоцисты, глазки и гонады. Радиальные каналы мы сюда не относим, так как это не самостоятельные отдельные органы, а лишь выпячивания желудка. Впрочем, вполне оставлять их без внимания не следует, так как гонады большею частью развиваются в ясной корреляции с радиальными каналами.
В наименьшем масштабе варьирует количество гонад. Так, у медуз Athecata (Anthomedusae), где гонады лежат на ротовом хоботке, число их колеблется между 4—6—8. У медуз Thecaphora (Leptomedusae) их чаще всего 4 или 8, у Trachymedusae наблюдается приблизительно то же самое, у Narcomedusae число отдельных скоплений половых клеток может быть большим и достигать такого количества, как щупальца (у Pegantha 10—30 щупалец и столько же половых скоплений клеток).
Глазки и статоцисты варьируют в числе от 4 и до нескольких сотен, но меньше четырех их, повидимому, не бывает.
Наконец, наиболее сильно способно изменяться число щупалец: от одного и до нескольких сот (рис. 3, в). Разобраться в отмеченном нами разнообразии необычайно трудно. Повидимому, большинство перечисленных органов могло во время эволюции медуз меняться в обе стороны, т. е. и в сторону олигомеризации и в направлении полимеризации.
Судя по распространенности четверного и восьмерного расположения гомологичных образований, одно из этих чисел и язляется основным у медуз. Является ли оно вместе с тем и наиболее примитивным, сказать гораздо более трудно, тем более, что по отношению к щупальцам мы можем в ряде случаев утверждать, что имела место вторичная олигомеризация их числа. Я говорю о тех медузах, у которых число щупалец ниже четырех, хотя вся остальная организация животного сохраняет четырех-лучевый характер. А таких примеров, как мы вскоре увидим, немало.
Особенная многочисленность случаев, когда щупалец 8 или 4, быть может, зависит от того, что при таком числе их вся периферия зонтика может достаточно хорошо и эффективно облавлиниться щупальцевым аппаратом — своего рода физиологический оптимум. Быть может, это обстоятельство вторично отозвалось и на числе радиальных каналов, а связь числа гонад ,с числом каналов не подлежит сомнению.
Имеется, однако, и ряд случаев дальнейшего уменьшения числа щупалец (рис. 2).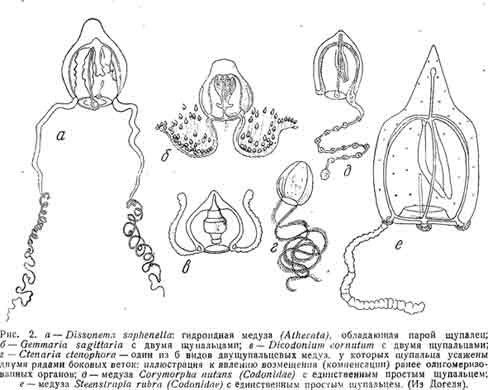
Среди медуз, относящихся к Athecata, двумя краевыми щупальцами обладают Thamnostylus, Cubogaster, Ctenaria, Gemmaria, Slomotoca, Amphinema, Dicodonitim, Zanclea, Perigonimus, миниатюрная Mnestra, паразитирующая на теле голожаберного планктонного моллюска Phyllirkoe.
Всего одно щупальце сохраняется у Corymorpha (Steenstrupia), Etiphysa, Hybocodon, Amphicodon, Microcampa, Heterostep Li anus.
Однако эта тенденция к редукции числа щупалец до 2 и 1 свойственна, по-видимому, только медузам Athecata.
Относительно Thecaphora и Trachylida соответственных данных я не нашел, если не считать двущупальцевую Hydroctena, которую сближают с Narcomedusae.
Зато у медуз Thecaphora мы находим один несомненный пример полимеризации—медузу Gastroblasta (рис 3, а), у которой И течение жизни умножается (до десятка) число ротовых хоботков и ртов, хотя число гонад остается равным четырем. Ланг считает, что в данном случае мы имеем перед собой не доведенное до конца бесполое размножение этой медузы делением.
В связи с олигомеризацией числа щупалец у многих медуз можно подметить одно чрезвычайно интересное явление, которое Не менее ясно выступает далее и у сифонофор, а именно: при олигомеризации щупалец сохраняющиеся щупальца получают тенденцию к ветвлению.
Иногда это ветвление является гребенчатым, т. е. всё веточки расположены друг за другом в один ряд; В других случаях оно перистое, наконец, в третьих — сама ось щупальца расщепляется на конце на 2—3 ветви, причем в отдельных случаях (Hybocodon, рис. 3, б) расщепление в базальцом направлении идет так далеко, что фактически получаются 2—3’отдельных щупальца, сидящих лишь на общем бугровидном основании.

Рис. 3. а—медуза из группы Thecaphora: Gastroblasta с полимеризацией ротовых хоботков и ртов; б — медуза Hybocodon (Amphicodon), у которой единственное щупальце расщепляется от основания на три отдельных щупальца на общем утолщенном основании: пример явления возмещения утраченных вследствие олигомеризации щупалец; в — Limnocnida tanganjicae — многощупальцевый вид трахимедуз. (Из Догеля).
Вопрос о такой вторичной „полимеризации» настолько интересен, что его стоит рассмотреть подробнее, имея в руках точные цифры. Для этого мы использовали многотомную сводку по медузам всего света, составленную Майером („Medusae of the World», 1910).
По этой сводке можно составить себе представление о точном числе видов медуз, имеющих различное число щупалец, и о тех соотношениях, которые имеются между количеством щупалец и степенью их ветвистости. Приводимая ниже таблица взята по данным Майера и охватывает собою всех гидроидных медуз, считая и Trachylida.
Распределение щупалец у различных медуз
| Число видов | |||||
| Число щупалец | всего | с неветвистыми
щупальцами |
то же | с ветвистыми
щупальцами |
то же |
| в процентах | в процентах | ||||
| 1 щупальце . . . | 13 | 13 | 100 | _ | _ |
| Я щупальца . . . | 37 | 32 | 86,5 | 5 | 13,5 |
| 4 щупальца . . . | 78 | 75 | 96 | 3 | 4 |
| 8 щупалец …. | 40 | 39 | 97,5 | 1 | 2,5 |
| более 8 щупалец | 328 | 328 | 100 | — | — |
| Всего . . . | 496 | 487 | 98 | 9 | 2 |
Но у этого вида непосредственно по бокам от основного щупальца на более поздних возрастных стадиях вырастает еще по щупальцу, вследствие чего билатеральная симметрия этих медуз, которые ранее обладали четырьмя радиально расположенными щупальцами, еще более Подчеркивается.
Первоначальное четырехщупальцевое строение однощупальцевых медуз хорошо явствует из того, что у некоторых из них еще сохранились очень маленькие рудименты 1—3 остальных щупалец.
Особенный интерес представляют двущупальцевые медузы. Среди 37 двущупальцевых гидромедуз 5, т. е. 13,5% обладают ветвистыми щупальцами (см. ранее сказанное).
Кроме этих 5 типично ветвистощупальцевых форм, мы находим у Eucodonium minerva, представителя двущупальцевых медуз, тенденцию к образованию ряда ветвей на обоих щупальцах. На эксумбреллярной стороне щупалец данного вида имеется правильный ряд довольно высоких бугров вроде зачатков щупалец.
Таким образом, к пяти указанным выше видам можно, собственно, прибавить еще один; в общем получится 6 видов, образующих от 37 около 16%.
Среди четырехщупальцевых медуз тоже имеется 3 вида с типичным гребенчатым ветвлением щупалец. Как видно, количество ветвистощупальцевых медуз среди видов, обладающих четырьмя щупальцами, значительно менее велико, чем среди двущупальцевых: всего 4%.
Еще менее велико оно у восьмищупальцевых медуз, у которых среди 40 таких видов медуз всего у одного, т. е. у 2,5%, имеются ветвистые щупальца. У медуз, число щупалец у которых превышает 8, разветвленные щупальца вообще отсутствуют.
Итак, можно прежде всего сделать вывод, что у медуз, обладающих 2—4—8 щупальцами изредка наблюдается тенденция к ветвлению щупалец, затухающая по направлению от двущупальцевых к восьмищупальцевым.
Мы исключаем из рассмотрения некоторых гидромедуз из сем. Cladonemadae (род Eleutheria). Считается, что у этих видов происходит дихотомия щупалец. Однако на самом деле легко видеть, что мы имеем здесь дело с тем же однорядным ветвлением, с той лишь разницей, что у Eleutheria образуется всего одна ветвь, и притом свободный конец самого щупальца дифференцируется в особый прикрепительный диск, или присоску. Одна из таких форм Eleutheria обладает 8 щупальцами (Е. dichotoma).
Что здесь имеет место ответвление, а не бифуркация щупалец, видно из следующего:
1) ветвь с прикрепительным диском составляет прямое продолжение щупальца, а другая ветвь торчит в сторону;
2) ветвь с прикрепительным диском иногда длиннее другой, „стрекательной» ветви;
3) „стрекательная» ветвь снабжена на конце булавой с книдоцистами, подобно многочисленным ветвям ранее рассмотренных однорядно ветвистых медуз;
4) „стрекательная» ветвь расположена по отношению к прикрепительной экстраумбреллярно, подобно ветвям на гребенчатых щупальцах.
. Таким образом, мы имеем достаточное основание считать, что прикрепительная „ветвь» с диском представляет на самом деле продолжение оси самого щупальца, тогда как „стрекательная» — единственное ответвление этой оси.
Наклонность к разветвлению щупалец настолько распространена у двущупальцевых медузоидных организмов, что представляет собою род закономерности, базирующейся на каких-то биологических или физиологических причинах (см. также сказанное далее о щупальцах у сифонофор и гребневиков).
Одна из этих причин, нам думается, поддается объяснению, если мы учтем, что ветвистость щупалец чаще всего встречается именно у двущупальцевых (гидромедузы, гребневики) и одно-щупальцевых (сифонофоры) форм, гораздо реже у четырехщупальцевых и в виде редчайших исключений у восьмищупальцевых форм.
Обычно же (кроме одного исключения) все восьмищупальцевые и все без исключения многощупальцевые медузы обладают простыми, неветвистыми щупальцами.
Очевидно, что олигомеризация числа щупалец до 2 или до 1 выходит за те пределы, которые являются оптимальными для вида, ибо такое падение их числа невыгодно отражается на способности животных к захвату добычи.
Соответственно с этим подобные виды развивают на щупальцах боковые или концевые ветви, чем увеличивается ловчая способность щупальца. Фактически у таких форм происходит возврат к многощупальцевому состоянию, совершающийся (согласно с правилом Долло), однако, новым способом.
Это явление, которое встретится нам и далее, можно назвать явлением возмещения, или компенсации утраченных вследствие олигомеризации органов.
Остается неразрешенным вопрос, почему не ветвятся, в таком случае, щупальца однощупальцевых Codonidae (систематика повсюду взята по Майеру).
Ответить на него мы можем только предположительно. Либо отсутствие компенсации у части Codonidae объясняется тем, что у части представителей этого семейства срок свободной жизни медузы после отрывания ее от полипоидной колонии особенно короток, почему сильное развитие органов захвата пищи не представляет необходимости.
При этом имеется один факт, говорящий в пользу такого соображения. Среди однощупальцевых медуз Codonidae имеется единственный вид, способный к бесполому размножению посредством отпочковывания молодых медузок от основания своего щупальца. Отсюда можно сделать вывод, что данный вид (Hybocodon prolifer) обладает достаточно длинным сроком планктонной жизни.
И как раз у этого вида по бокам его основного щупальца на более поздних стадиях роста вырастают в непосредственном соседстве с основанием щупальца еще два щупальца; впрочем, может быть, их правильнее было бы называть побочными ветвями единственного главного щупальца, достигшими известной самостоятельности.
Во всяком случае у Н. ргоlifer, тем или иным способом, происходит компенсация двух щупалец из трех, исчезнувших до степени маленьких рудиментов щупалец.
Кроме того, следует припомнить, что у всех сифонофор, т. е. форм, гастрозоиды которых обладают единственным щупальцем, последнее всегда разветвлено.
Вас также заинтересует это:
Comments (0)
Комментариев нет
Нет комментариев.