05.01.2016
Интенсивность и ритм дыхания трепанга
Рубрика: ИглокожиеАвтор: admin
Сведения об интенсивности потребления кислорода дальневосточным трепангом немногочисленны.
Ю. Э. Брегман (1973) исследовал этот показатель стандартным методом замкнутых сосудов и с помощью его модификации, позволившей проследить этот процесс в динамике.
Использование такой методики позволило установить, что вслед за начальным значительным повышением потребления кислорода наступает относительная стабилизация обмена.
По мере снижения концентрации кислорода в камере его потребление постоянно, хотя и медленно, снижалось. Для расчета использовали как средине данные, полученные в модифицированных камерах, так и результаты опытов в обычных закрытых камерах.
В опытах Ю. Э. Брегмана потребление кислорода дальневосточным трепангом составило 0,03 мл/г-ч. По его данным, рассматриваемый вид способен адаптировать скорость потребления кислорода при изменении температурных условий среды обитания, что объясняется эвритермностью трепанга. Вместе с тем имеются сведения (Choe, 1963), что интенсивность потребления кислорода этой голотурией зависит от температуры.
С. Чои сообщил, что с повышением температуры воды у дальневосточного трепанга быстро возрастал уровень потребления кислорода (рис. 29), а величина степенного показателя в уравнении обмена снижалась. По мнению Ю. Э. Брегмана (1973), подобное быстрое изменение параметров уравнения обмена приблизительно в среднем интервале температур (13,5—18,5°С) представляется маловероятным. Брегман полагает, что на результаты Чои повлияли какие-то ненормальные условия проведения опытов.
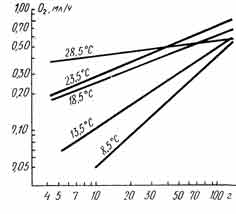
Рис. 29. Зависимость интенсивности потребления кислорода дальневосточным трепангом с различной массой кожно-мышечного мешка от температуры. Шкалы по осям логарифмические
В этой связи значительный интерес представляют результаты, полученные А. В. Жакиным (в печати). Он проводил измерения потребления кислорода дальневосточным трепангом с февраля по ноябрь в среднем ежемесячно. Голотурий (масса кожно-мышечного мешка 70—90 г) собирали в зал. Посьета с глубины 3—4 м. Эксперименты проводили в море непосредственно в месте расположения садков с животными.
Потребление кислорода в этих опытах в течение года колебалось от 0,0017+0,0004 мл/г-ч (март, t = 0,5°C) до 0,0127 =0,0011 мл/г-ч (начало августа, t = 26,0°C). В тех же пределах лежат значения потребления кислорода, которые получил С. Чои (Choe, 1963), однако диапазон колебаний величины рассматриваемого показателя, полученный им, несколько уже. Это, несомненно, связано с тем, что А. В. Жакин использовал в опытах животных, физиологическое состояние которых менялось в разные сезоны.
В годовом ходе интенсивности обмена у дальневосточного трепанга А. В. Жакин выделяет зоны зимнего минимума, весенне-летнего максимума, зону толерантности и осеннего спада. Зимний минимум интенсивности обмена легко объяснить низкими температурами воды.
Увеличение обмена в период весенне-летнего максимума связано с повышением температуры воды и с интенсивным ростом гонад. Именно развитием генеративных органов можно объяснить значительную разницу в величинах обмена в весенний и осенний периоды при одинаковых значениях температуры. Конец периода весеннего максимума совпадает с массовым нерестом голотурий в начале августа.
Зона толерантности приходится на конец августа — конец сентября. Величина потребления кислорода в этот период остается относительно постоянной при значительных колебаниях температуры (15,2—23,0°С) и близка к скорости потребления кислорода при 9—8°С. Уменьшение обмена в августе — сентябре можно связать с потерей животными части биомассы с половыми продуктами и внутренними органами при сезонной эвисцерации.
В октябре при температуре 15—11С отмечена некоторая интенсификация обмена. Она обусловлена переходом животных к активному образу жизни после эстивации, ростом гонад и восстановлением внутренних органов. Последующий период характеризуется постепенным снижением обмена вслед за уменьшением температуры воды до зимнего уровня.
Таким образом, зависимость интенсивности потребления кислорода от температуры в значительной степени определяется физиологическим состоянием голотурий и может меняться в разные периоды года.
Для исследования ритма дыхания дальневосточного трепанга и измерения объема прокачиваемой через клоаку воды было сконструировано специальное устройство (Левин, Гочаков, 1979).
Устройство (рис. 30) состоит из колпачка, который с помощью эластичной мембраны с отверстием укрепляется на анальном конце тела голотурии.
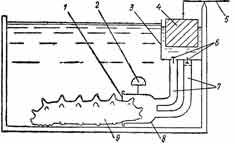
Рис. 30. Устройство для регистрации интенсивности дыхания голотурий. 1—мембрана с отверстием; 2 — компенсирующий поплавок; 3 — поплавковая камера; 4— регистрирующий поплавок; 5 — указатель; 6 — клапаны; 7 — гибкие трубки; 8— колпачок; 9 — экспериментальное животное
Двумя шлангами, снабженными направленными в противоположные стороны клапанами, колпачок связан с камерой, в которую помещен регистрирующий поплавок. Для компенсации веса колпачка он снабжен компенсационным поплавком.
В отличие от аппаратов, предложенных для этой цели ранее (Newell, Courtney, 1965; Robertson, 1972), разработанное нами устройство почти не препятствует движению голотурий и позволяет получить характеристики ритма и «глубины» дыхания в условиях, приблиратуры.
При температуре 19—20°С для трепанга длиной тела около 20 см объем прокачиваемой за один «вдох» воды составляет 4—6% общего объема тела. Эта величина почти не отличается от полученной Д. Робертсоном (Robertson, 1972) для S. mollis.
Зная количество кислорода, которое организм голотурии усваивает с помощью водных легких, и количество прокачиваемой через них воды с известным содержанием кислорода, несложно определить эффективность поглощения кислорода через стенки легких.
Эта величина у дальневосточного трепанга составляет 30—50%, что несколько ниже определенной Д. Робертсоном у S. mollis. Относительно невысокая эффективность поглощения кислорода водными легкими объясняется примитивностью функциональных механизмов, обеспечивающих газообмен — медленной и плохо организованной циркуляцией омывающей водные легкие перивисцеральной жидкости и отсутствием тесного контакта терминальных пузырьков левого легкого со стенками кровеносных сосудов «чудесной сети».
Несмотря па значительное участие водных легких в газообмене при нормальном состоянии животного, потеря этих образований при эвисцерации мало сказывается на дыхании дальневосточного трепанга.
После экспериментальной эвисцерации ритм клоакальной пульсации сохраняется почти неизменным, хотя при дыхательных движениях вода через разрыв стенки клоаки поступает непосредственно в полость тела.
При этом, несмотря на отсутствие водных легких, скорость потребления кислорода организмом практически не уменьшается. Это, несомненно, следует связать с непосредственным поступлением морской воды к внутренней поверхности стенки тела.
В то же время лишение внутренностей сказывается на повышении чувствительности организма дальневосточного трепанга к, понижению содержания кислорода в воде, что отмечено и для S. mollis (Dimock, 1977).
Вас также заинтересует это:
Comments (0)
Комментариев нет
Нет комментариев.