19.12.2015
ТРАГАКАНТОВЫЙ ТИП СОКРАЩЕНИЯ КОРНЕЙ РАСТЕНИЙ
Рубрика: ФлораАвтор: admin
Г. Д. Ярошенко (Получено 27.11.1945)
Различными авторами описано явление сокращения корней, наблюдаемое у многих многолетних растений, которое вызывает втягивание под поверхность почвы нижних частей стеблей этих растений, углубление в глубину почвы луковиц, корневищ и т. д.
Первое обстоятельное исследование в этой области принадлежит Гуго де-Фризу, который установил явление сокращения корней у Cynam scolymus, Verbascum. ihapsus и других растений.
По данным де-Фриза, это объясняется увеличением тургора в клетках паренхимы, в особенности в более молодых тканях, вблизи камбиального слоя.
При этом клетки паренхимы корней увеличиваются в объеме, а весь корень растягивается в поперечном направлении, укорачиваясь одновременно по длине.
Daniel [3], Rimbach [6] изучали явление сокращения корней у однодольных растений, благодаря которому луковицы этих растений (Colchicum, Allium, Crocus, Gladiolus, £Lilium и др.) с каждым годом втягиваются в почву на все большую глубину.
Семена этих растений прорастают, как известно, на поверхности почвы, но через несколько лет луковицы оказываются погруженными в землю на глубину до 20 см и даже более.
По Daniel, укорачивание корней однодольных объясняется сокращением объема отдельных клеток паренхимы, вследствие потребления растением находившихся в них запасов питательных веществ.
Rimbach объясняет явление сокращения корней у Lilium и других однодольных увеличением объема клеток на периферии корня при одновременном сокращении объема клеток паренхимы в центральной части корней. При этом увеличения диаметра корней не происходит.
Ducelier описал явление сокращения корней у Oxalis cernua, вызывающее постепенное погружение в глубину почвы подземных клубней этого растения. Thoday описал аналогичное явление у Oxalis incamata и Brodiaea lactea.
По Thoday сокращение корней связано с уменьшением, а не с увеличением их диаметра и объясняется отмиранием и сокращением объема части клеток паренхимы, после отдачи ими запасов питательных веществ и воды.
Последнее является приспособительной реакцией к условиям засухи, позволяющей растениям безболезненно переносить периоды засухи. При сокращении корней кора их покрывается горизонтальными кольцевыми складками.
Church [2] подвергла критике работы De Vries и Rimbach; она сообщает, что у двудольных сокращение корней вызывает искривление сосудистых пучков, в то время как у однодольных сосудистые пучки сохраняют прямолинейное направление.
Нами изучен новый тип сокращения корней различных растений, наиболее резко выраженный у трагакантовых астрагалов, вследствие чего нами и предлагается для подобных случаев термин «трагакантовый тип» сокращения корней.
Описанные до сих пор случаи сокращения корней возникают в результате изменения объема клеток паренхимы корней.
Трагакантовый же тип сокращения корней вызывается размножением клеток паренхимы сердцевины и сердцевинных лучей корней. Это разрастание паренхимы корней не связано с деятельностью камбия, происходит даже в старых тканях, сосредоточенных в центральной части корня, возраста (например в случае трагакантовых астрагалов) в несколько десятков лет, и объясняется тем, что паренхима сердцевины и сердцевинных лучей сохраняет мерисгематический характер в течение всей жизни растений.
Сокращение корней трагакантовых астрагалов изучено нами у произрастающих в Армянской ССР видов Astragalus microcephalus W., A. strictifofius Boiss. и др. [‘].
У трагакантовых астрагалов процесс разрастания паренхимы сердцевины и сердцевинных лучей протекает следующим образом. В течение всей жизни куста астрагала, на протяжении 70 и более лет, происходит постепенное расширение полости центральной сердцевины как в корне растения, так и в стволе и в ветвях.
Обычно у древесно-кустарниковых пород сердцевина образуется в течение первого года жизни растения и в дальнейшем больше не растет. Дальнейший рост в толщину корня, ствола и ветвей происходит путем отложения камбием годичных колец на периферии корня и ствола. У трагакантовых же астрагалов наблюдается постепенное расщирение сердцевинной полости.
У молодых экземпляров диаметр сердцевинной полости равен 1.0—1.5 мм. У старых кустов сердцевинная полость расширяется до диаметра 0.4—1.0 см, а в отдельных случаях и выше. Это расширение сердцевинной полости объясняется постепенным разрастанием паренхимы, распирающей стенки сердцевинной полости и вызывающей этим увеличение ее диаметра.
Процесс этот связан с образованием трагакантовой камеди при распаде клеточных стенок паренхимы сердцевины. Распадаются клетки в центральной части сердцевины, в то время как на периферии расширенной полости сердцевины сохраняется все время тонкий слой живой размножающейся паренхимы.
В корнях и стволах старых растений вся сердцевинная полость представляется заполненной камедью.
Тонкий слой паренхимы у стенок центральной сердцевинной полости заметен лишь при микроскопическом исследовании. Распадаются с образованием камеди, по всей вероятности под влиянием специального энзима, только стенки клеток паренхимы, окрашивающейся хлор-цинк-иодом в синий цвет. Одеревеневшие стенки сосудов и либриформа, окрашивающиеся хлор-цинкиодом в красно-оранжевый цвет, не распадаются.
Аналогичный процесс разрастания паренхимы наблюдается также в сердцевинных лучах и также связан с образованием и накоплением у них трагакантовой камеди. Полости сердцевинных лучей у старых кустов также представляются сплошь заполненными камедью; тонкий слой живой паренхимы находится лишь у стенок полостей. Разрастание паренхимных лучей происходит в тангентальом направлении.
Между отдельными сердцевинными лучами ткани корня и ствола представлены главным образом сосудами и либриформом, длина клеток которых остается неизменной. Поэтому длина периферии тангентального разреза сердцевинного луча также остается постоянной.
Увеличение плоскостей тангентальных сечений сердцевинных лучей при разрастании паренхимы достигается путем изменения щелевидной формы тангентальных сечений в округлую, что сопровождается уменьшением размеров их в вертикальном направлении, при одновременном увеличении размеров их в тангентальном направлении.
Это явление иллюстрируется рис. 1 и 2, на которых представлены в тангентальном сечении молодая древесина корня Astragalus microcephalus (рис.1) с нерасширенными еще площадями сечений сердцевинных лучей и старая древесина того же растения, в центральных годичных слоях (рис. 2), с расширенными в горизонтальном направлении площадями тангентальных сечений сердцевинных лучей.
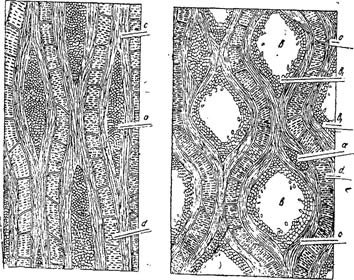 Рис. 1. Fig. 1. Тангентальный разрез молодой древесины корня Astragalus mlcrocephalas
Рис. 1. Fig. 1. Тангентальный разрез молодой древесины корня Astragalus mlcrocephalas
а — сердцевинные лучи с неразложившейся паренхимой, с—либриформ, d — сосуды
Рис. 2. Fig. 2. Тангентальный разрез старой древесины корня Astragalus microcephalus.
а —сердцевинные лучи с неразложившейся паренхимой,
b — заполненные камедью лолосги сердцевинных лучей на местах разложившейся паренхимы,
b-1 — живая паренхима, выстилающая -стенки полостей сердцевинных лучеq,
с — либриформ,
d — сосуды
Это явление иллюстрируется бумажной моделью, представленной на рис. 4, схематизирующей тангентальный разрез древесины корня астрагала. При растягивании модели в горизонтальном направлении площади сечений прорезанных в бумаге щелей увеличиваются, что сопровождается сокращением размеров фигуры в вертикальном направлении и увеличением их в горизонтальном направлении.
На корнях растений это явление сопровождается сокращением длины корня при увеличении его диаметра. Последнее происходит независимо от деятельности камбия; таким образом, увеличение с возрастом диаметра корней у трагакантовых астрагалов происходит двумя путями:
1) вследствие деятельности камбия, откладывающего на периферии древесинного цилиндра годичные слои, и
2) благодаря независимому от этого процесса непрерывному размножению клеток паренхимы сердцевины и сердцевинных лучей.
В результате сокращения корней шейка пня корней с возрастом постепенно втягивается под поверхность почвы; у кустов возраста 35—40 лет шейка пня бывает углублена обычно под поверхность почвы на 15—20 см. Окружающие сердцевинные лучи сосудисто-волокнистые пучки в молодой древесине астрагалов имеют почти вертикальное направлений (рис. 1).
В старой древесине сосудистые пучки искривлены (рис. 2). В корнях старых кустов астрагалов наружные, т. е. наиболее молодые годичные слои, имеют древесину с щелевидными, тангентальными разрезами сердцевинных лучей и мало искривленными сосудистыми пучками.
По мере приближения к центру корня или ствола сердцевинные лучи все более расширяются, а сосудисто-волокнистые пучки все более искривляются, что наиболее резко выражено в древесине наиболее старых центральных годичных слоев.

Рис. 3. Fig. 3. Тангентальный разрез через годичные слои различного возраста«одного и того же корня Astragalus microcepfiilus возраста 35 лет.
а — наружные годичные слои возраста 3—4 лет,
b — годичные слои возраста 15—20 лет,
с — самые старые центральные годичные слон возраста 30—35 лет
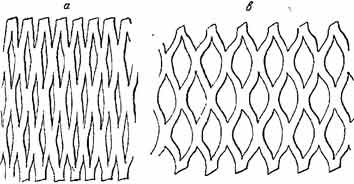 Рис 4. Fig. 4. Бумажная модель, иллюстрирующая разрастание паренхимы cердцевинных лучей у трагакантовых астрагалов, приводящее к сокращению корней.
Рис 4. Fig. 4. Бумажная модель, иллюстрирующая разрастание паренхимы cердцевинных лучей у трагакантовых астрагалов, приводящее к сокращению корней.
а — в нерастянутом виде, b —в растянутом виде
Это явление представлено на рис. 3, на котором изображены тангентальные сечения древесины корней одного и того же куста Astragalus microcephalus, взятые в годичных слоях разного возраста. По мере образования трагакантовой камеди, последняя, поглощая воду и набухая, создает давление, расширяющее еще более полости сердцевины и сердцевинных лучей и усиливающее явление сокращения корней.
В старой древесине трагакантовых астрагалов сосудисто-волокнистые пучки настолько искривлены, что на тангентальных разрезах создают подобие сетки, в петлях которой находятся полости сердцевинных лучей (рис. 3, с).
Такую структуру древесины мы предлагаем называть «трагакантовой структурой» тканей древесины. Эта структура чрезвычайно характерна для данного типа сокращения корней, обнаруженного нами и у ряда других растений.
«Трагакантовый тип» сокращения корней очень распространен в природе и обнаружен нами у целого ряда травянистых многолетников, часто весьма далеко отстоящих в систематическом отношении как. друг от друга, так и от рода Astragalus.
Однако процесс сокращения корней у этих растений, хотя и принадлежит в общем трагакантовому типу, т. е. обусловливается независимым от деятельности камбия размножением клеток паренхимы, но у разных растений представляет некоторое различие, что позволяет выделить несколько вариантов данного процесса, а именно:
 Риг. 5. Fig. 5. Тангентальный разрез корня f lumbago europaea возраста 7 лет.
Риг. 5. Fig. 5. Тангентальный разрез корня f lumbago europaea возраста 7 лет.
а — периферический годовой слой возраста 2 лет, b — внутренний годичный сдой возраста 5 лет. Увелич. 1X2
Рис. 6. Fig. 6. Тангентальный разрез корня Symphytum caucasicum. Увелич. 1 X 1,6
1) Процесс расширения с возрастом тканей тангентальных сечений сердцевинных лучей и связанный с ним процесс сокращения корней протекает в точности по типу трагакантовых астрагалов.
Эта картина наблюдается у травянистых многолетних представителей рода Astragalus: A. brachycarpus MB., A. Stevenianus и др. Далее, тот же тип обнаружен нами у Medicago sativa, Medicago coerulea, Plumbago europaea, Statice Gmelini, Centaurea Glehnii, Cetilaurea Beketi, Centaurea diffusa, Echinops Szovitsii, Kochia prostrata, Gypsophila bicolor’.
У Astragalus brachycarpus разрастание паренхимы сопровождается распадом ее с образованием трагакантовой камеди как в сердцевине, так и в сердцевинных лучах. У Centaurea Glehnii клеточные стенки паренхимы сердцевины также распадаются с образованием камеди, повидимому не отличающейся по свойствам от камеди трагакантовых астрагалов.
У всех остальных исследованных нами растений этой группы распада паренхимы вовсе не обнаружено. Здесь разрастающаяся паренхима полностью заполняет и расширяет полости сердцевины и сердцевинных лучей, вызывая этим сокращение корней.
У Plumbago europaea (рис. 5) характерно преобладание паренхимы над элементами сосудисто-волокнистых пучков в периферических частях старых корней. Это, однако, объясняется не более интенсивным разрастанием паренхимы в периферических годичных слоях, а своеобразной деятельностью камбия, откладывающего с увеличением возраста растения все больше и больше паренхимы по сравнению с клетками сосудисто-волокнистых пучков.
Эта паренхима дает начало вторичным сердцевинным лучам, не доходящим до центра корня. При этом типе сокращения корней шейка пня бывает втянута в глубину почвенного слоя относительно глубоко, например у старых многолетников возраста 7—10 лет на глубину до 10—20 см.
2) Второй вариант представлен случаями, когда главная масса корней образована паренхимой, среди которой вертикально протянуты тяжи сосудисто-волокнистых пучков; иногда, как у Serratula quinqtiefolia, эти пучки не соприкасаются друг с другом.
В таком случае сердцевинные лучи не обособлены и ясно не выражены, как, например, у Symphytum caucasicum (рис. 6).
В других случаях, при общем преобладании в тканях корней паренхимы, относительно удаленные друг от друга тонкие сосудисто-волокнистые пучки образуют правильную сеть, подобно тому, как это наблюдается у трагакантовых астрагалов.

Рис. 7. Fig. 7. Тангентальный разрез корня Telekia. speciosa. a. —центральная часть девятилетнего корня, b — периферическая часть молодого корня возраста 2 лет. Увелич. 1 х 1,4
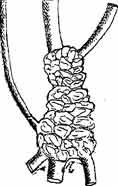 Рис. 8. Fig. 8. Узловатое корневище Psephellus somcheticus 4/5 н. в.
Рис. 8. Fig. 8. Узловатое корневище Psephellus somcheticus 4/5 н. в.
Такая структура корней характерна, например, для Telekia speciosa (рис. 7). К этой группе относятся Serratula quinquefolia, Symphytum caucasicum, Telekia speciosa,Cephalaria giganiea. В старых тканях сосудисто-волокнистые пучки волнообразно искривлены (рис. 7), что и указывает на происшедшее сокращение корней по длине. Шейка пня у названных растений возраста 7—10 лет углублена под поверхностью почвы до 10—15 см.
3) Третий вариант относится к растениям, имеющим короткое узловатое корневище с отходящими от него корнями (рис. 8). Процесс сокращения при наличии трагакантовой структуры резко выражен только в узловатом корневище и не выражен или выражен лишь в незначительной степени в корнях (рис. 9).
Таким образом, шейка пня у этих растений втягивается в почву, повидимому, лишь в результате сокращения узловатого корневища, в связи с чем при этом варианте шейка пня бывает погружена в почву сравнительно неглубоко, в пределах примерно 3—5 см. Из обследованных нами растений сюда относятся Psephellus somcheticus и Lathyrus roseus.
4) Наконец, у ряда растений трагакантовая структура тканей корней резко выражена только в лубе и слабо выражена в древесине корней. Сюда относятся из исследованных нами растений Otiobrychis transcaucasica (рис. 10) и Astragalus glycyphylloides. В данном случае процесс сокращения корней, повидимому, протекает различно в древесине и в лубе. Этот интересный вариант подробно изучен нами не был и нуждается в дополнительных исследованиях.
Ни в одном из случаев вариантов 2—4 распада паренхимы с образованием камеди нами не обнаружено.
Трагакантовый тип сокращения корней может быть определен по следующим признакам:
1) Шейка пня, т. е. граница стебля и корня, всегда понижена под поверхностью почвы, в то время как у ряда растущих растений других видов шейка пня находится нормально у поверхности почвы.
2) Сосудисто-волокнистые пучки сильно искривлены на тангентальных разрезах, в особенности в более старых (центральных) слоях корня. На тангентальных разрезах корней обычно ясно бывает выражена «трагакантовая структура».
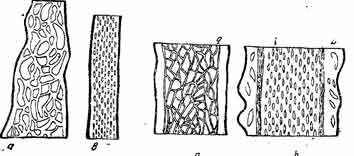 Рис. 9. Fig. 9. Тангентальные разрезы узловатого корневища (а) и корня (6) Psephellas somcheticus. Увелич. 1 X 1.6
Рис. 9. Fig. 9. Тангентальные разрезы узловатого корневища (а) и корня (6) Psephellas somcheticus. Увелич. 1 X 1.6
Рис. 10. Fig. 10. Тангентальний разрез древесины (а) и луба (й) корня Onobrvchis transcaucasica. Увелич. 1 X 3,2
Последняя бывает наиболее выражена в более старых по возрасту (центральных) тканях корней.

Рис. 11. Fig. 11. Полости в сердцевине центрального старого корня Telekia speciosa. В полостях видны оставшиеся клочья мертвой паренхимы. Радиальный разрез. Увелич. 1X2
Рис. 12. Fig. 12. Сокращение корней у Telekia speciosa, вследствие роста в толщину боковых корней
Рис. 13. Fig.13. Сокращение корней у Pyrethrum myriophyllam, вызванное спиральным завертыванием корней
3) Площади тангентальных сечений сердцевинных лучей в центральных частях корня заметно превышают таковые в периферических частях корней. Увеличение площади сечений достигается расширением полости сердцевинных лучей в горизонтальном направлении при одновременном сокращении их длины в вертикальном направлении. Форма сечений изменяется при этом из щелевидной в округлую.
4) Среднее число клеток паренхимы на тангентальных разрезах сердцевинных лучей в старых (центральных) тканях корня (с расширенными площадями сердцевинных лучей) значительно превышает таковое в молодых (периферических) тканях при приблизительно одинаковых размерах клеток.
Собственно процесс разделения клеток сердцевинной паренхимы, т. е. явления кариокинеза, нами ни в одном случае замечены не были. По-видимому, этот процесс ограничен во времени и протекает лишь в определенные сезоны и определенные часы суток, уловить которые нам не удалось, тем более, что большинство растений нами было исследовано осенью, когда процессы роста замедляются или прекращаются.
У трагакантовых астрагалов, а также у некоторых других растений, у которых разрастание паренхимы связано с ее распадом и образованием камеди, весь процесс является приспособительной реакцией к условиям засухи, так как накопляющиеся запасы трагакантовой камеди, относящейся к категории так называемых гидрофильных коллоидов, служат аккумулятором влаги и этим обусловливают высокую степень ксерофильности этих растений.
В большинстве же случаев, когда разрастание паренхимы и сердцевинных лучей не связано с образованием камеди, трагакантовый тип сокращения корней различных растений наблюдается как у ксерофитов, так и у влаголюбов, произрастающих даже и в условиях избыточной влажности (как, например, у Telekia speciosa).
У ряда растений, например у Telekia speciosa и Centaurea diffusa, в старых корнях паренхима сердцевины и сердцевинных лучей отмирает, но без образования камеди; в местах отмирания паренхимы образуются полости (рис. 11).
К трагакантовому типу сокращения корней примыкает другой тип сокращения корней, вызываемый ростом в толщину боковых корней, ответвляющихся от главного корня. При росте в толщину боковых корней увеличивается в диаметре основание их, проходящее через ткани центрального корня, что вызывает такое же раздвигание и искривление окружающих их сосудисто-волокнистых пучков; отсюда и укорочение центрального корня (рис. 12).
В небольшой степени этот тип сокращения корней проявляется почти у всех многолетников, у которых боковые корни отходят от верхней части центрального корня. Более резко сокращение центрального корня при росте в толщину боковых корней проявляется тогда, когда оно соединяется с трагакантовым типом сокращения корней.
Наконец у Pyrethrum myriophyllum старые корни распадаются на отдельные деревянистые тяжи, весь пучок которых завертывается спирально (рис. 13), что в связи с ростом боковых корней также приводит к сокращению корней и погружению шейки пня в почву на глубину до 5—8 см.
ЛИТЕРАТУРА
[1]Аствацатрян З. и Ярошекко Г., К биологии трагакантовых астрагалов Арм. ССР, Известия Арм. фил. Акад. Наук СССР, №1, 1941,—[2] Church М., Root contraction, The Plant World (Tuczon, Arizona), v. 22, No. 11, 1919,— [3] D a-nlelL., Sur les racines napiformes, transitoldes des Monocotyledones.Rev.Gen.de Bot., p. 455, 1891.—[4] DeVries Hugo, Ueoer die Konlraction der Wurzeln, Landw: Jahrbuch., IX, 1, p. 37, 1880.—[5] Ducelier M. L„ Note sur la vegetation de VOxalis cernua, Rev. Gen. de Bot., XXV, 2, p. 217, 1914.—[6] Rimbach A., Bio-logische Beobachtungen an Colchicam autumnale, Ber. deutsch. Bot. Ges., XV, 298; 12, 1897 и др.—[7] Thoday D., Tht contractile Roots of Oxalis incarnata, Annales of Botany, London, v. XL, No. CLIX, 1926.— [8] Thoday D„ Mechanism of root contraction in Brodiaea lactea, Plant Physiology, 6, 721—725, 1931.
Ботанический институт
Академии Наук Армянской ССР
Вас также заинтересует это:
Comments (0)
Комментариев нет
Нет комментариев.