25.07.2015
Сложности, возникающие при клонировании в плазмидах больших фрагментов ДНК
Рубрика: МикромирАвтор: admin
Соотношение трансформантов, содержащих рекомбинантные плазмиды, и трансформантов с повторно замкнутым в кольцо вектором может зависеть также от размера встраиваемой чужеродной ДНК.
Как правило, чем больше вставка чужеродной ДНК, тем ниже эффективность трансформации. Поэтому при клонировании больших фрагментов ДНК (> 10 kb) особенно важно принять все возможные меры для снижения повторного замыкания в кольцо молекул вектора.
Но и в этом случае фон оказывается довольно высоким, и обычно для выявления рекомбинантных трансформантов необходимо использовать гибридизацию in situ (Grunstein, Hogness, 1975; Hanahan, Meselson, 1980).
БАКТЕРИОФАГ λ (лямбда)
С тех пор как впервые было показано, что бактериофаг λ можно использовать в качестве вектора (Murray, Murray, 1974; Rambach, Tiollais, 1974; Thomas et al., 1974), на его основе сконструировано большое число разнообразных векторов (см. обзор Williams, Blattner, 1980).
Чтобы эффективно использовать эти векторы, необходимо знать основы молекулярной биологии бактериофага λ (более исчерпывающую информацию см. в работах Herskowitz, 1973; Hendrix et al., 1982).
Бактериофаг λ представляет собой вирус, содержащий двух-цепочечную ДНК. Его геном имеет размер около 50 kb. ДНК в частицах бактериофага λ находится в виде линейных двухцепочечных молекул с одноцепочечными комплементарными концами длиной по 12 нуклеотидов (липкие концы).
Вскоре после проникновения в бактерию-хозяина эти концы слипаются, ДНК замыкается в кольцо и в течение ранней стадии инфекции транскрибируется уже как кольцевая молекула.
В ходе этой стадии происходит выбор одного из двух альтернативных путей репликации вируса.
1. При литическом развитии кольцевая ДНК многократно реплицируется в клетке, синтезируются большие количества продуктов генов бактериофага, образуются и созревают частицы вирусного потомства.
В конце концов клетка лизируется, высвобождая много новых инфекционных вирусных частиц.
2. При логенизном же развитии геном инфицирующего бактериофага встраивается в ДНК бактерии хозяина, последовательно реплицируется и передается клеткам-потомкам как обычный хромосомный ген.
Ниже подробнее описаны те гены и их продукты, которые участвуют в этих двух путях развития бактериофага λ.
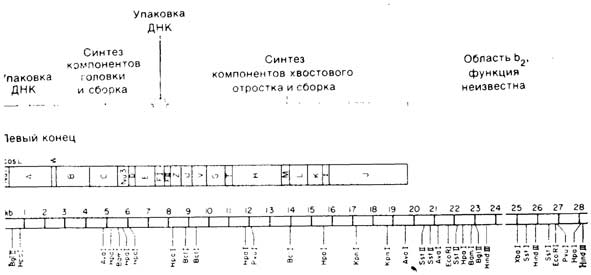
Рис. 1.5. Физическая и генетическая карты бактериофага лямбда дикого типа. Ввертического и лизогенного путей развития. Далее приведена генетическая карта, функции каждого из этих генов см. в работе Hendrix et al., 1982). Внизу три-молекулы в тысячах пар оснований (kb). Отмечены сайты расщепления рес-работе Уильямса и Блаттнера (Williams, Blattner, 1980).
Литический цикл
Немедленная ранняя транскрипция
В начале инфекции транскрипция инициируется на двух промоторах, рl и рr которые расположены непосредственно слева и справа от гена сI (кодирующего репрессор) (рис. 1.7). Терминация получающихся при этом «немедленных ранних» РНК происходит в конце генов N и cro соответственно в сайтах и Tl TR1 хотя рост некоторых правосторонних транскриптов продолжается и далее, через гены О и Р (которые кодируют белки, участвующие в репликации ДНК) и заканчивается в сайте Левосторонний транскрипт кодирует белок N, действие которого необходимо для следующей стадии инфекции.
Задержанная ранняя транскрипция
Белок N нейтрализует активность фактора терминации р клетки-хозяина и позволяет транскрипции пройти через ранние терминаторы tL, и tr1 в остальную область ранних генов. Поэтому белок N является положительным регуляторным элементом, активность которого необходима для литического развитии бактериофага, обладающего сайтом tr2.
Мутанты могут, однако, расти (хотя и относительно плохо), если в результате делеции у них удален сайт tr2. Такие фаги называют мутантами nin (N-independent — N независимыми).
Репликация ДНК
В течение ранней стадии инфекции кольцевая ДНК фага лямбда реплицируется двунаправленно в виде структур Кэрнса (или тета-форм). Эта репликация начинается в единственной точке инициации репликации (ori) (рис. 1.7), для активации которой необходимы два кодируемых вирусом белка — О и Р.
Позднее начинается репликация по типу катящегося кольца (чем определяется переход от структур Кэрнса к катящемуся кольцу — неизвестно) и образуются катенированные линейные молекулы ДНК.
В процессе упаковки ДНК продукт гена А фага лямбда расщепляет эти катенаты в сайтах cosL и cosR, в результате чего получаются те линейные молекулы ДНК единичной длины с липкими концами, которые и оказываются в зрелых частицах бактериофага.
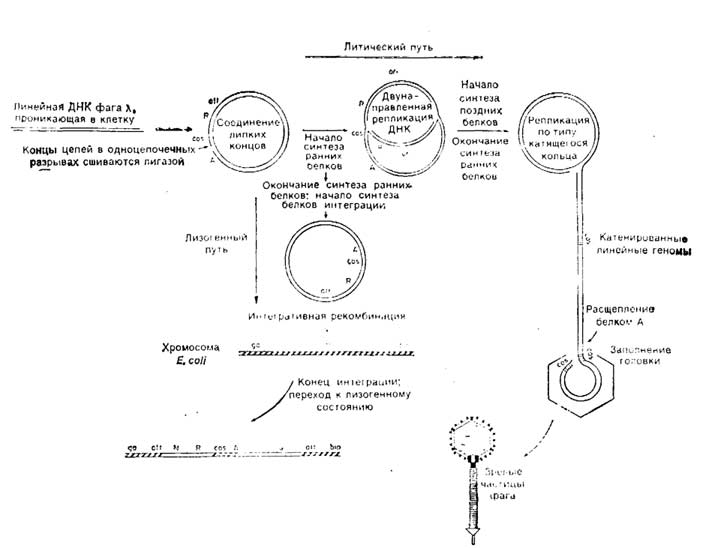 Рис. 1.6. Схема жизненного цикла бактериофага лямбда. Проникающая в бактерию-хозяина линейная двухцепочечная ДНК фага X изображена двойной линией.
Рис. 1.6. Схема жизненного цикла бактериофага лямбда. Проникающая в бактерию-хозяина линейная двухцепочечная ДНК фага X изображена двойной линией.
Для литического пути развития, который указан горизонтальной стрелкой, необходимо функционирование как ранних, так и поздних генов. Для лизогенного же пути, который указан вертикальной стрелкой, нужны функции лишь ранних генов.
К этому времени, однако, образуется другой регуляторный белок — Q, причем в количествах, достаточных для того, чтобы началась транскрипция поздних генов.
Белок Q действует как положи-тельный регулятор синтеза РНК с промотора р\, который используется для транскрипции всей поздней области, содержащей многочисленные гены, ответственные за сборку головки и хвостового отростка и за лизис клетки.
Сборка
Два основных компонента зрелых частиц — головка и хвостовой отросток — собираются по отдельности.
Самым ранним из выявленных при сборке головки предшественником является «обнесенная лесами» предголовка (рис. 1.8). Дальнейшее созревание предголовок, в ходе которого происходят удаление белка, выполняющего функцию «лесов», и протеолитическое расщепление других компонентов, зависит от функционирования белка groE бактерия-хозяина.
В результате получаются структуры, названные предголовками. На первом этапе упаковки ДНК в предголовку участвуют два белка, Nul и А, которые связываются с катенированной линейной ДНК вблизи левого сайта cos, но не с самим этим сайтом.
Затем такой комплекс присоединяется к определенной области предголовки. В присутствии белка FI ДНК укладывается внутри предголовки, размер которой при этом увеличивается примерно на 20%. Когда головка оказывается заполненной, снаружи капсида присоединяется белок D («decoration» — белок «наружной отделки»), который «обжимает» головку вокруг ДНК.
В процессе упаковки левый и правый сайты cos на катенированной линейной ДНК оказываются рядом у входа в головку, где они и расщепляются наискосок под действием ter-функции белка А с образованием липких концов длиной по 12 нуклеотидов. Наконец, заполненные головки объединяются с независимо образованными хвостовыми отростками.
Лизис
Для лизиса клетки-хозяина и высвобождения потомства фага необходимы два фаговых белка, R и S. Особенно часто используются мутанты по белку S, так как при этом внутри клеток накапливаются большие количества фаговых частиц и тем самым повышается выход фага. Мутанты S~ можно высвободить, лизируя клетки хлороформом.
Вас также заинтересует это:
Comments (0)
Комментариев нет
Нет комментариев.